
A few years ago during a monitoring trip to Provincetown, I took a tour of the 250 ft Pilgrim Monument (350 ft above the harbor) and climbed to the top to discover an outstanding view of the marina and harbor that gave a different perspective on the monitoring site. The outer harbor has a deep channel 50 to 70 ft deep that curves around the west end of the Cape (Lands End), making Provincetown one of the deepest marine fishing harbors along the north Atlantic coast. The inner harbor is formed by a 2,500 foot long stone breakwater built by the Army Corps of Engineers in the early 1970's about 835 ft off the end of the Provincetown Wharf complex.
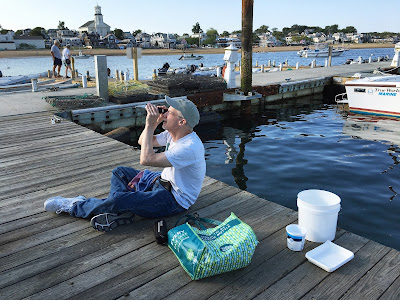
Ascidians
During the 2015-2017 summer seasons, the docks were inspected for invasive species in the MIMIC program (Coast Watch 2011, Coast Watch 2012). The most abundant attached species were the colonial ascidians Botryllus, Botrylloides, Didemnum, and Diplosoma, and the solitary ascidian Styela. The one exception to the general abundance of ascidians was the rare ocurence of the solitary species Ascidiella aspersa, which has been periodically seen at low frequency in Provincetown (several years only 1 specimen), but not seen in 2017. Interestingly, the translucent solitary species Ciona intestinalis and Molgula manhattensis, cryptogenic species not included in the survey have also been observed at low frequency whereas the colonial species and the solitary Styela are common all over the marina. Perhaps these three translucent species are out-competed and crowded out by the colonial species which spread throughout the marina in the summer. Interestingly, Molgula manhattensis is abundant in Wellfleet marina, a location with very low species diversity and few colonial species of any kind.
The rock shrimp Paleomon is easiest seen in late summer when they have grown larger than Paleomonetes shrimp and the blue bands on their legs are prominently displayed. Unlike the other species in the program, Paleomon requires an active search for individuals by running a net along the sides of the floating docks where algae and hydroids are numerous and closely inspecting the shrimp for morphological features.
Algae
The Asian red alga Grateloupia has been a consistent member of the floating dock community albeit at relatively low frequency in comparison to other invasives. It is usually found on the same docks where Codium is located near the pilings that hold the docks in place. An analysis of of the MIMIC data presented on the MORIS website for the distribution of Grateloupia along the MA/NH/ME coast indicates that it has not yet been reported yet at MIMIC sites north of Boston and has remained in southern New England since it was first observed in the Gulf of Maine in 2007. Other MA sites where it has been observed include the east end of the Cape Cod Canal at Sandwich, Onset and New Bedford on Buzzards Bay, and Fall River on the upper Naragansett Bay.
Codium fragile on Floating Dock

Grateloupia turuturu on Floating Dock

A cluster of Grateloupia turuturu growing on a floating dock right below the water line, Provincetown, August 2017.
Bryozoans
During the last 2 years, the bryozoan Bugula neritina has remained a common species on the docks along with a newly identified invasive bryozoan Tricellaria inopinata. The bryozoan Tricellaria was first identified in Massachusetts in 2010 in Eel Pond, Woods Hole, lower Cape Cod (Johnson et al, 2012) and was reported in the Gulf of Maine during the 2013 Rapid Assessment Survey (RAS), where it was given the common name "unexpected bryozoan". I have seen Tricellaria since I started monitoring (unidentified beige bryozoan in my November 2012 Bugula neritina post (MIS Branching Bryozoan), but I only became aware that it was an invasive species in 2016. On the floating docks, Bugula and Tricellaria grow in the same niche and frequently cohabitate together. Last July, the colonies were typically only a few cm high and newly settled juveniles were common. Tricellaria has a distinctly different microscopic growth pattern from other native, beige Bugula species (zooids point toward the anterior whereas they are alternately opposed to each other in Bugula). This pattern could be discerned in the lab under a dissecting microscope and can be seen at the docks with a 20x or 30x magnifying lens. During the summer of 2017, Tricellaria was particularly abundant and overgrew other species like the green algae Ulva and Codium.
In contrast to the branching bryozoa, the encrusting bryozoan Membranipora membranacea has not been seen much in Provincetown despite the expectation that it might be seen on a floating dock. The rare times it has been seen has been on kelp growing on the dock or plastic dock trim that was submerged in the water. The marina does not seem conducive to other encrusting bryozoans either, since the native encrusting broyzoans Electra pilosa and Schizoporella unicornis, are also rarely seen.
 Purple Bugula and light beige Tricellaria growing with green alga Ulva and orange chain tunicate Botrylloides, August, 2017. Recently settled juvenile Tricellaria can be seen growing on the Ulva surfaces.
Purple Bugula and light beige Tricellaria growing with green alga Ulva and orange chain tunicate Botrylloides, August, 2017. Recently settled juvenile Tricellaria can be seen growing on the Ulva surfaces.

 Tricellaria and Neosiphonia growing on Codium in an area of docks where Codium and Grateloupia are common, August 2017. Heavy growth on the base of Codium obscurs the holdfast and lower branches.
Tricellaria and Neosiphonia growing on Codium in an area of docks where Codium and Grateloupia are common, August 2017. Heavy growth on the base of Codium obscurs the holdfast and lower branches.
View of Provincetown Harbor from Pilgrim Monument

View of Provincetown Harbor and Wharf from the Pilgrim Monument. MacMillan Pier on the left, Fisherman's Wharf on the right. The stone breakbeakwater is seen beyond the end of the piers. Truro is seen a few miles across the harbor in the far background. The tip of Lands End is shown on the right.
View of MacMillan Pier from Pilgrim Monument
 MacMillan Pier showing the docks, fishing vessels, and tour boats. View from the Pilgrim Monument looking south. On the east side (left), two floating dock complexes are closest to shore followed by two finger piers near the end of the main pier. On the west side (right), a long row of tour boats line the pier. The ferry terminal to Boston is located at the end of the pier on the west side (right).
MacMillan Pier showing the docks, fishing vessels, and tour boats. View from the Pilgrim Monument looking south. On the east side (left), two floating dock complexes are closest to shore followed by two finger piers near the end of the main pier. On the west side (right), a long row of tour boats line the pier. The ferry terminal to Boston is located at the end of the pier on the west side (right).
Both floating dock complexes are monitored each July and August for invasive species. The salinity and temperature are recorded and photographs are taken to document salient features. Salinity is typically fairly high and ranged from 31 to 35 parts per thousand between 2012 and 2017. Temperatures ranged from 22 to 27 degrees C (approximately 70 to 80 degrees F).
Measuring Salinity on the North Floating Dock

Holding a refractometer toward the light to make a reading. Ready for monitoring with my checklist, thermometer, refractometer, camera, jeweler's loupes for magnifying, buckets, shallow pan for documentation, and small net for capturing crustaceans.
Ascidians
During the 2015-2017 summer seasons, the docks were inspected for invasive species in the MIMIC program (Coast Watch 2011, Coast Watch 2012). The most abundant attached species were the colonial ascidians Botryllus, Botrylloides, Didemnum, and Diplosoma, and the solitary ascidian Styela. The one exception to the general abundance of ascidians was the rare ocurence of the solitary species Ascidiella aspersa, which has been periodically seen at low frequency in Provincetown (several years only 1 specimen), but not seen in 2017. Interestingly, the translucent solitary species Ciona intestinalis and Molgula manhattensis, cryptogenic species not included in the survey have also been observed at low frequency whereas the colonial species and the solitary Styela are common all over the marina. Perhaps these three translucent species are out-competed and crowded out by the colonial species which spread throughout the marina in the summer. Interestingly, Molgula manhattensis is abundant in Wellfleet marina, a location with very low species diversity and few colonial species of any kind.
Crustaceans
The skeleton shrimp Caprella mutica and the European rock shrimp Paleomon elegans were seen with low frequency. The skeleton shrimp prefers hydroids and algae like Ulva in the more protected waters
near the pier and has been consistently observed since its disappearance after Hurricane Irene in August 2011.
The rock shrimp Paleomon is easiest seen in late summer when they have grown larger than Paleomonetes shrimp and the blue bands on their legs are prominently displayed. Unlike the other species in the program, Paleomon requires an active search for individuals by running a net along the sides of the floating docks where algae and hydroids are numerous and closely inspecting the shrimp for morphological features.
Algae
The Asian red alga Grateloupia has been a consistent member of the floating dock community albeit at relatively low frequency in comparison to other invasives. It is usually found on the same docks where Codium is located near the pilings that hold the docks in place. An analysis of of the MIMIC data presented on the MORIS website for the distribution of Grateloupia along the MA/NH/ME coast indicates that it has not yet been reported yet at MIMIC sites north of Boston and has remained in southern New England since it was first observed in the Gulf of Maine in 2007. Other MA sites where it has been observed include the east end of the Cape Cod Canal at Sandwich, Onset and New Bedford on Buzzards Bay, and Fall River on the upper Naragansett Bay.
Codium fragile on Floating Dock

A large clump of Codium fragile growing at the waterline on a polystyrene float under one of the secondary docks, Provincetown, July, 2016. The attachment site is clearly shown and there are no epiphytes growing on its surface.
Grateloupia turuturu on Floating Dock

During the last 2 years, the bryozoan Bugula neritina has remained a common species on the docks along with a newly identified invasive bryozoan Tricellaria inopinata. The bryozoan Tricellaria was first identified in Massachusetts in 2010 in Eel Pond, Woods Hole, lower Cape Cod (Johnson et al, 2012) and was reported in the Gulf of Maine during the 2013 Rapid Assessment Survey (RAS), where it was given the common name "unexpected bryozoan". I have seen Tricellaria since I started monitoring (unidentified beige bryozoan in my November 2012 Bugula neritina post (MIS Branching Bryozoan), but I only became aware that it was an invasive species in 2016. On the floating docks, Bugula and Tricellaria grow in the same niche and frequently cohabitate together. Last July, the colonies were typically only a few cm high and newly settled juveniles were common. Tricellaria has a distinctly different microscopic growth pattern from other native, beige Bugula species (zooids point toward the anterior whereas they are alternately opposed to each other in Bugula). This pattern could be discerned in the lab under a dissecting microscope and can be seen at the docks with a 20x or 30x magnifying lens. During the summer of 2017, Tricellaria was particularly abundant and overgrew other species like the green algae Ulva and Codium.
In contrast to the branching bryozoa, the encrusting bryozoan Membranipora membranacea has not been seen much in Provincetown despite the expectation that it might be seen on a floating dock. The rare times it has been seen has been on kelp growing on the dock or plastic dock trim that was submerged in the water. The marina does not seem conducive to other encrusting bryozoans either, since the native encrusting broyzoans Electra pilosa and Schizoporella unicornis, are also rarely seen.
Bugula and Tricellaria Growing on Dock Float and Algae

Closer View of Bugula and Tricellaria

A clump of Bugula and Tricellaria growing out from the dock float on a mass of undergrowth. The notched branches of Bugula, which represent alternating left and right facing zooids, can be discerned at the outer edges of the bush.
Tricellaria Overgowth on Codium

LINKS:
Marine Invasive Species in the MIMIC Program: Coast Watch 2011, Coast Watch 2012
Mathieson, AC, Dawes, CJ, Pederson, J, Gladych, RA, and Carlton, JT. The Asian red seaweed Grateloupia turuturu (Rhodophyta) invades the Gulf of Maine. Biol. Invasions 10: 985-988, 2008.
Johnson, CH, Winston, JE, and Woollacott, RM. Western Atlantic introduction and persistence of the marine byrozoan Tricellaria inopinata. Aquatic Invasions 7: 295-303, 2012.
Mathieson, AC, Dawes, CJ, Pederson, J, Gladych, RA, and Carlton, JT. The Asian red seaweed Grateloupia turuturu (Rhodophyta) invades the Gulf of Maine. Biol. Invasions 10: 985-988, 2008.
Johnson, CH, Winston, JE, and Woollacott, RM. Western Atlantic introduction and persistence of the marine byrozoan Tricellaria inopinata. Aquatic Invasions 7: 295-303, 2012.
RAS Report: Report on the 2013 Rapid Assessment Survey of Marine Species at New England Bays and Harbors.
MORIS: CZM's Online Mapping Tool.
MORIS: CZM's Online Mapping Tool.
Tricellaria Sighting in Provincetown, 2012: MIS Branching Bryozoan






































